
Translate this page into:
Hepatokines and Adipokines in Metabolic Syndrome

Address for correspondence SatinathMukhopadhyay, Department of Endocrinology & Metabolism, Institute of Post-Graduate Medical Education & Research-Seth Sukhlal Karnani Memorial Hospital (IPGME&R-SSKM), Kolkata, India (e-mail: satinath.mukhopadhyay@gmail.com).
This article was originally published by Thieme Medical and Scientific Publishers Pvt. Ltd. and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Hepatokines and adipokines are secretory proteins derived from hepatocytes and adipocytes, respectively. These proteins play a main role in the pathogenesis of metabolic syndrome (MetS), characterized by obesity, dysglycemia, insulin resistance, dyslipidemia, and hypertension. Adipose tissue and liver are important endocrine organs because they regulate metabolic homeostasis as well as inflammation because they secrete adipokines and hepatokines, respectively. These adipokines and hepatokines communicate their action through different autocrine, paracrine and endocrine pathways. Liver regulates systemic homeostasis and also glucose and lipid metabolism through hepatokines. Dysregulation of hepatokines can lead to progression toward MetS, type 2 diabetes (T2D), inflammation, hypertension, and other diseases. Obesity is now a worldwide epidemic. Increasing cases of obesity and obesity-associated metabolic syndrome has brought the focus on understanding the biology of adipocytes and the mechanisms occurring in adipose tissue of obese individuals. A lot of facts are now available on adipose tissue as well. Adipose tissue is now given the status of an endocrine organ. Recent evidence indicates that obesity contributes to systemic metabolic dysfunction. Adipose tissue plays a significant role in systemic metabolism by communicating with other central and peripheral organs via the production and secretion of a group of proteins known as adipokines. Adipokine levels regulate metabolic state of our body and are potent enough to have a direct impact upon energy homeostasis and systemic metabolism. Dysregulation of adipokines contribute to obesity, T2D, hypertension and several other pathological changes in various organs. This makes characterization of hepatokines and adipokines extremely important to understand the pathogenesis of MetS. Hepatokines such as fetuin-A and leukocyte cell-derived chemotaxin 2, and adipokines such as resistin, leptin, TNF-α, and adiponectin are some of the most studied proteins and they can modulate the manifestations of MetS. Detailed insight into the function and mechanism of these adipokines and hepatokines in the pathogenesis of MetS can show the path for devising better preventative and therapeutic strategies against this present-day pandemic.
Keywords
metabolic syndrome
hepatokine
adipokine
type 2 diabetes
obesity
hypertension

Introduction
Metabolic syndrome (MetS) is a collection of related health issues that has a close association with T2D and cardiovascular diseases. It has turned into an epidemic disorder and describes many clinical and metabolic risk markers. MetS comprises abdominal adiposity and dyslipidemia characterized by insulin resistance, inflammation, low high density lipoprotein (HDL) cholesterol, high triglycerides, and blood pressure, which also correlate with cardiovascular disease (CVD).1,2
MetS has been defined in various aspects by the World Health Organization (WHO). Other groups working on epidemiology, endocrinology, basic science, public health have also proposed their definition on MetS.3,4,5 In the present day definition, MetS has been framed according to NCEP-ATP III and other clinical and biochemical parameters.
Studies have shown a varying prevalence of MetS in people above the age of 20 years, ranging from around 20% to 39.3% in India, the USA, Tehran, Turkey and Saudi Arabia.6,7 MetS is becoming more common due to an increase in the occurrence of obesity and may overtake smoking as the main cause behind heart disease in future.
The major risk markers of metabolic syndrome are described below and mentioned in ►Fig. 1.
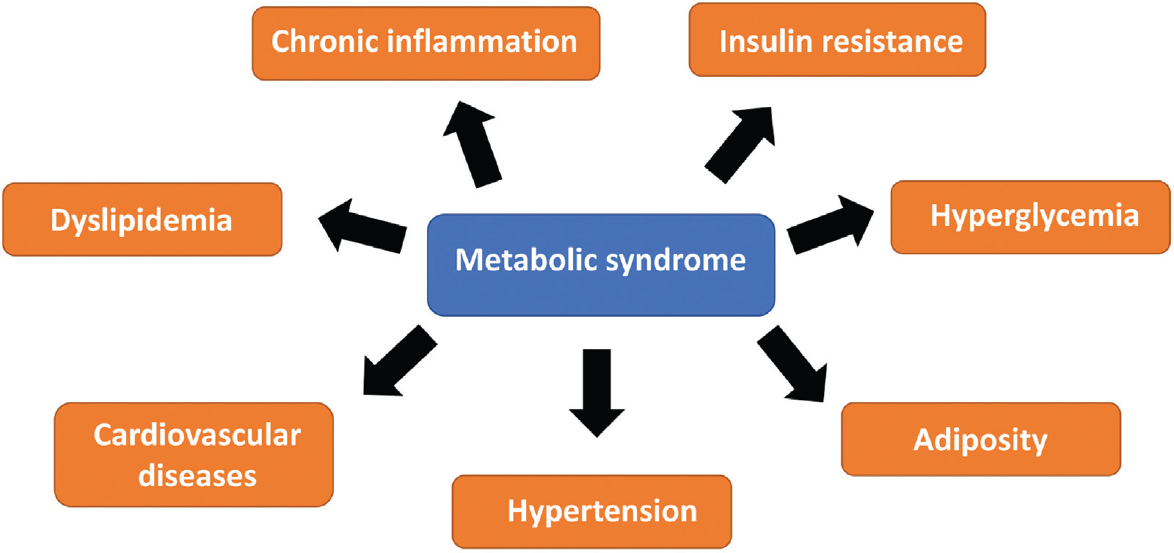
- Major risk markers of metabolic syndrome.
1. Adiposity
Being overweight and deposition of abdominal fat is the root cause of MetS. Body fat deposits in different regions in females and males following separate patterns. These tendencies of fat deposition are called gynoid and android adiposity because women tend to deposit excess adipose tissue in outlying regions like legs, and pelvic region while men have tendencies to deposit adipose tissue in the central region (like abdomen). However, the position of adipose tissue is very complex because the regions of body fat deposition are common in men and women. Notably, deposition of adipocytes in abdominal region creates more complexities and lead to MetS. It leads to glucose intolerance, where the action of insulin is shunted.8
2. Insulin resistance
Insulin resistance (IR) is a key factor in MetS, linking the other components stated above. These metabolic abnormalities are the outcomes of the interaction between glucose utilization and storage in skeletal muscles and adipose tissue, culminating in increased insulin demand and hyperglycemia. Healthy women and men have fat content between 18% and 20% and 10 and 15% of body weight, respectively. MetS affects 30 to 40% of people, mainly due to an increase in weight, and intra-abdominal or ectopic fat accumulation. Reducing weight by 5 to 7% by diet and workout can lower MetS, risk of type 2 diabetes, and cardiovascular diseases. Moreover, anti-diabetic drugs that can improve insulin resistance, reduce the level of lipid and control weight gain are helpful for people with type 2 diabetes arising out of MetS. Bariatric surgery is another option for those with body mass index (BMI) greater than or at least 40 kg/m2 or 35 to 40 kg/m2 with significant co-morbidities.9
3. Type 2 Diabetes (T2D)
The possibility of T2D increases by five times in MetS patients. MetS and insulin resistance together have a cumulative effect because these patients are more prone to develop T2D. T2D is often linked to chronic inflammation. MetS in females, who have pregnancy-induced diabetes, also significantly escalates the likelihood of developing T2D.10
4. Dyslipidemia
In insulin-sensitive individuals, insulin inhibits adipose tissue lipoprotein lipase. Insulin resistance therefore enhances lipolysis in adipose tissue, resulting in FFA overproduction, increasing its plasma level and also uptake by hepatocytes. FFA increases liver triglyceride and cholesterol ester content. High level of triglycerides in blood TG content induces function of cholesterol ester transfer protein (CETP), that in turn stimulates removal of triglyceride from very low-density lipoprotein (VLDL) and adding to high-density lipoprotein (HDL), hence HDL clearance increases. It also stimulates the conversion of TG into LDL. LDL again in turn undergoes lysis, resulting in the formation of tiny and concentrated cholesterol less LDL particles, which increases the risk of cardiovascular diseases.11
5. Hypertension
Hypertension has become a global pandemic, developing countries facing the brunt of the problem, where the number of hypertensives are estimated to increase by 80% by 2025.12 Approximately 1 billion people are affected globally by hypertension and associated heart and kidney disease. It is the one of the major causes behind global mortality. Hypertension is intricately linked to insulin resistance, high cholesterol, cardiovascular diseases, and MetS. Hyperinsulinemia or the increased insulin levels promote hypertension as a compensation for lesser insulin-mediated glucose uptake by skeletal myocytes. Moreover, resistin, and leptin secretion from adipocyte increases during hypertension and insulin resistance.13,14
Adipokines in Metabolic Syndrome
Adipokines have a wide variety of role in multiple physiological functions leading to abnormal characteristics of MetS. Anomalous production of adipokines from visceral fat contribute to a proinflammatory condition. An interplay of low adiponectin and high leptin and resistin contribute to dyslipidemia, hyperinsulinemia, and atherosclerosis. To date, only leptin has been developed into a drug.15,16 Dipeptidyl peptidase-4 (DPP-4) has been recognised as an adipokine and DPP-IV inhibitors (gliptins) are already in use for the treatment of type 2 diabetes.17,18 Extensive research is needed to identify the basic mechanism of action of adipokines such as apelin, chemerin and resistin. Deficiency in understanding the mechanism of action of these adipokines has retarded the development of adipokine-based therapeutic strategies.19 Further research is needed to identify the specific adipokines with potential diagnostic and therapeutic properties. The role of different adipokines in MetS are discussed below and shown in ►Fig. 2.

- Adipokines in metabolic syndrome: role of resistin, IL-6, TNF-α, adiponectin, and leptin in dyslipidemia, insulin resistance, inflammation, type 2 diabetes, and hypertension.
1. Resistin
The most crucial point about resistin is that it counteracts insulin's action. Its expression is obstructed during adiposity as well as in T2D. This inhibition of the expression of resistin is mediated by increased expression of TNF-α. Under normal condition, resistin particularly decreases the repressive action of insulin so that gluconeogenesis can occur in the liver. When resistin does not function, circulatory glucose concentrations decreases and insulin sensitivity improves. The gene expression and secretion of resistin increases during adipogenesis in 3T3-L1, which in turn inhibits 3T3-LI adipogenesis.20,21 Circulatory resistin remains high in obese and T2D patients.22 However, in certain cases, resistin levels remain quite low in patients having adiposity as well as insulin resistance, which makes the role of resistin controversial. Hyperglycemia is caused by excess secretion of resistin due to increased hepatic glucose production. Resistin also subdues the activation of AMP-activated protein kinase (AMPK) in hepatocytes and muscle cells.23,24 However, these restrictive actions of resistin upon the activation of AMPK do not occur in primary culture of murine muscle cells, advocating that this process requires release of other factors from other cell types.25 Moreover, the level of resistin in circulation increases with increasing BMI in humans as well as rodents.24 A recent meta-analysis showed circulatory resistin levels to be higher in hypertensive subjects compared to their normotensive counterparts, suggesting a pathogenic role of resistin in hypertension.26 Nevertheless, additional investigation is necessary to identify the detailed function of resistin in MetS.
2. Interleukin-6 (IL-6)
Interleukin-6 (IL-6) is considered as a very important adipocytokine that regulates immune responses. It is produced by fibroblasts, monocytes, and adipocytes. Adipocyte-derived IL-6 makes up almost 15 to 35% of total circulating IL-6.27 The receptor of IL-6 functions by regulating other signaling molecules.28 It must be mentioned that plasma concentrations and adipocyte-derived IL-6 levels rise during obesity.28 In high-fat diet (HFD) fed mice, elevated levels of IL-6 in adipocytes induce insulin resistance, inhibiting the function of insulin receptor via proteasomal degradation of insulin receptor substrate (IRS).29 A key component related with IL-6 is C-reactive protein, and their concentrations are positively correlated.30 The presence of IL-6 is found in a wide range in different fat or lipid deposition sites. For example, IL-6 levels in visceral adipocytes are higher than peripheral depositions. Altogether, the relation between IL-6 concentration and T2D is quite complicated. Level of IL-6 in circulation increases in obese and T2D patients with significantly enhanced concentrations in visceral adipocytes.31 IL-6 activates AMPK-activated protein kinase in myotubes; however, the detailed signaling pathway involved in IL-6-mediated glucose uptake and other downstream elements in AMPK activation are not known.32 Hence, IL-6, a pro-inflammatory cytokine, can be regarded as an indicator of insulin resistance, adiposity, and T2D.33
3. Tumor Necrosis Factor-alpha (TNF-α)
Just like IL-6, TNF-α is also highly expressed in visceral adipocytes than peripheral adipocytes. It is profusely released by macrophages. TNF-α levels in adipocytes increase with obesity, BMI, insulin resistance, and T2D.34 TNF-α lowers the level of resistin and also impairs its function in 3T3-L1 adipocytes. Blocking of TNF-α affects other adipokines and improves inflammatory markers. It must be mentioned that TNF-α impairs insulin function in liver and adipocytes.35 Incubation with TNF-α affects insulin-mediated glucose uptake in skeletal muscle cells. Moreover, TNF-α receptor-IgG chimeric protein restores phosphorylation and insulin signaling via its receptor and substrate (insulin receptor substrate-1/IRS-1) in adipocytes and skeletal muscle cells. Also, knockout of TNF-α or TNF-α receptor improves glucose homeostasis and increases insulin sensitivity in obese models but not in obese patients with T2D.36,37 In short, TNF-α levels in adipocytes is positively correlated with fasting plasma glucose levels, insulin, and triglyceride levels in high fat-fed female subjects. It also increases adipocyte lipolysis.38 Hence, TNF-α elevates insulin resistance because it stimulates the production of lipids from adipocytes. Macrophages that are produced from adipose tissue resident monocytes are another significant source of pro-inflammatory cytokines. Also, the ratio of pro-inflammatory and anti-inflammatory cytokines regulate insulin resistance, heart, and kidney diseases.39
4. Adiponectin
Adiponectin is a very crucial adipocyte-derived protein because it is a key regulator of obesity and T2D. While it can induce release of proinflammatory cytokines, it also has anti-inflammatory functions and affects nuclear factor (NF-kB). It also enhances the action of insulin in peripheral tissues. Adiponectin decreases glucose concentrations and induces liberation of fat without increasing insulin levels, hence acting as an endogenous insulin sensitizer. Adiponectin is circulation occurs in three forms: trimer, hexamer, and high molecular weight (HMW) 12-18-mer adiponectin.40 It enhances energy expenditure in hepatocytes and skeletal muscle cells, while reducing tissue triglyceride content. Adiponectin restores insulin sensitivity through AMPK-induced reduction of gluconeogenesis in the liver and increased glucose uptake by muscles.41 There are two major receptors for adiponectin-AdipoR1 and AdipoR2, having seven transmembrane domains. AdipoR1 is found in muscles, and AdipoR2 is found in the liver.42 Increased plasma level of adiponectin is known to significantly reduce the risk of T2D. High molecular weight (HMW) adiponectin is an important biomarker for MetS, and mainly responsible for increased insulin sensitivity.43 Hypoadiponectinemia is typical in metabolic and cardiovascular disease states such as T2D, non-alcoholic hepatic steatosis, hypertension, and heart disease. Genetic hypoadiponectinemia is mediated by a missense mutation. Decrease in adiponectin levels precede the progression of T2D, making it an important component behind the condition. Adiponectin levels affect the formation of glucose in the liver by decreasing gene expression of enzymes: phosphoenolpyruvate carboxykinase as well as glucose-6-phosphatase. These two enzymes are very important for gluconeogenesis. Adiponectin also aids in the formation of foam cells from macrophages and reduction of TNF-α production from macrophages.44 Adiponectin induces weight loss and ameliorates dyslipidemia. Adiponectin expression is reduced in people with metabolically unhealthy adiposity (diabesity).45 Increased adipocyte accumulation in the visceral region is linked to several health issues. Interestingly, individuals with higher visceral adipose tissue (VAT) have low level of circulatory adiponectin.46 Adiponectin manifests glucose tolerance by insulin-sensitivity, energy utilization, and anti-inflammatory functions. Adiponectin also aids in the development of MetS in women after menopause.47 A recent clinical study on post-menopausal women showed that a decrease in body weight significantly increased adiponectin levels and reduced insulin in circulation in fasting condition and also IR.48 The most cost-effective way of increasing adiponectin, the major anti-inflammatory adipokine, is reduction of the adipose tissue mass by achieving around 5 to 7% weight loss through hypocaloric diet and physical exercise.49,50,51 Also, recent studies have shown that GLP-1 agonists, i.e., semaglutide and liraglutide and dual GLP-1 and GIP agonist tirzepatide, can increase adiponectin and reduce pro-inflammatory cytokines, by inducing 10 to 20% weight loss.52,53 PPAR-gamma agonists such as pioglitazone and rosiglitazone significantly improve insulin sensitivity by increasing adiponectin levels and reducing fetuin-A levels, albeit with loss of weight.54,55
5. Leptin
Leptin is essentially a thinning hormone, inhibiting food intake by stimulating the hypothalamic satiety center. Leptin is secreted majorly from adipocytes. However, small quantities of leptin are secreted from skeletal muscle, liver, placenta, and fundus of the stomach. Leptin levels increase with increasing amount of fat deposition in body. High level of leptin is associated with obesity and T2D.56 Leptin resistance leads to unregulated food intake and often co-exists with hypothalamic insulin resistance. The various mechanisms by which a loss of hypothalamic leptin signal leads to weight gain are induction of leptin by Suppressor of Cytokine Signalling-3 (SOC-3), decreased circulation to brain, and increased endoplasmic reticulum (ER) stress. The JAK-STAT pathway activated by leptin receptor is also an important regulator of increasing BMI and phosphoinositol-3-kinase is also regulated by leptin, which, in turn, controls glucose metabolism.57 Circulatory leptin levels modulate nutritional status because its level decreases soon after fasting begins.58 Leptin secretion is also dependent on body fat content. Also, 80% of total leptin production is from subcutaneous fat. Circulatory leptin levels, and its secretion from adipose tissue are positively correlated with obesity. Higher content of fat is linked to an increase in leptin resistance. Leptin is a crucial moderator in managing body weight. Hence, deficiency of leptin in blood leads to leptin resistance. Leptin resistance is a key player behind adiposity.59 Leptin improves insulin sensitivity through AMPK. AMPK in turn decreases malonyl-CoA, thereby impairing acetyl-CoA carboxylase. This explains how leptin deficiency contributes to insulin resistance. It establishes that leptin resistance as a key factor behind adiposity and can be targeted to reduce insulin resistance. Bariatric surgery achieves these goals in a much more robust way, as it allows patients to achieve more than 20% weight loss. DiRECT trial, where participants were subjected to very low-calorie diets (600 calories), resulted in 12 to 15 kg weight loss.60,61,62,63 This was associated with significant improvement in obesity-associated insulin and leptin resistance, secondary to favorable improvements in adipo-hepatokine profiles. Although the actual part played by leptin in regulating insulin resistance is unclear, the contribution of leptin along with other adipokines such as TNF-α, resistin, IL-6 in MetS is confirmed.64 Finally, leptin is a major gate keeper of the initiation of puberty in girl. Adequate leptin levels tell the hypothalamic GnRH neurons that the body has acquired enough adipose tissue store to sustain pregnancy and child birth; the latter then triggers the onset of puberty.65
Hepatokines in Metabolic Syndrome
Liver, one of the major glands in human body, plays several functions such as regulating carbohydrate, fat, and protein metabolism. Hepatokines, the liver-derived secretory proteins, have been given the status of ‘new hormones’ having substantial role in MetS. The hepatokines regulate energy homeostasis by controlling glucose and lipid metabolism.66 The high prevalence of type 2 diabetes has increased the need for drug development targeting the liver. Many glucose-lowering drugs are used as hepatokine blockers for treating liver diseases. Some known drugs are nuclear hormone receptor agonists such as peroxisome proliferator-activated receptor agonists, and farnesoid X receptor agonists, incretins, fibroblast growth factor-19/21 and sodium–glucose cotransporter inhibitors.67 However, detailed studies on deciphering the mechanism is necessary to evaluate hepatokine induced tissue damage and the role of these drugs in metabolic syndrome. Several hepatokines have been identified that provide important knowledge in understanding the pathogenesis of MetS. These hepatokines pose to be the possible targets for treating obesity and other related diseases. The role of all these hepatokines in MetS are as follows and shown in ►Fig. 3:

- Hepatokines in metabolic syndrome: role of Hepatocyte-derived fibrinogen-related protein 1 (HFREP1) or Fgl1 or hepassocin, Fetuin-A, fibroblast growth factor 21 (FGF-21), angiopoietin-related growth factor (AGF), angiopoietin-related growth factor (AGF), angiopoietin-like proteins (ANGPTL) and leukocyte cell-derived chemotaxin 2 (LECT2) in dyslipidemia, insulin resistance, inflammation, type 2 diabetes, and hypertension.
1. Fetuin-A
Fetuin-A (AHSG), is a liver-derived protein and is one of the major culprits behind T2D via blocking the function of insulin receptor and promoting secretion of cytokines. Fetuin-A reduces adiponectin secretion, in adipocytes and monocytes, which is a main protein that plays important role against MetS. Fetuin-A secretion increases with increased hepatic fat accumulation and sub-clinical atherosclerosis while it is negatively associated with insulin sensitization.68 The position of fetuin-A gene is 3q27. Single nucleotide polymorphisms (SNPs) in human AHSG gene is linked to T2D.69 The role of excess fetuin-A is reported in obesity, MetS, and T2D. Fetuin-A can be a potential bio-marker for T2D.69 Interestingly, plasma fetuin-A levels can be reduced by calorie restriction.70 Pioglitazone, an insulin sensitizer, decreases fetuin-A mRNA expression in liver tissue.54 Regular exercise or workout has variety of action on circulatory fetuin-A levels; and it has been proven that exercise can reduce serum fetuin-A concentration in T2D while increasing serum adiponectin levels. Lowering fetuin-A level in obese subjects can lead to improvement in IR. Increased serum fetuin-A levels following weight loss after 6 months of aerobic training in healthy overweight old men has been shown to be cardioprotective.71 Altogether, fetuin-A is a very promising drug target for preventing and treating T2D.66
2. Hepatocyte-derived fibrinogen-related protein 1 (HFREP1)
Hepatocyte-derived fibrinogen-related protein 1 (HFREP1, or Fgl1 or hepassocin), is produced from the liver and can lead to progression of T2D through ERK1/2. HFREP1 is also secreted in adipocytes where it controls lipid homeostasis. Plasma HFREP1 levels correlate positively with insulin desensitization and the rate of progression of prediabetes to T2D.72
3. Fibroblast growth factor 21 (FGF-21)
FGF-21, another hepatokine, is a strong energy regulator and an essential hormone needed for adaptive starvation. FGF21 is also a myokine and an adipokine. It also affects the level of reactive oxygen species (ROS), ER stress, and other cellular processes and exerts a favorable effect on controlling body weight and triglyceride and cholesterol levels. FGF21 increases GLUT-1 expression in the skeletal muscle and insulin-mediated glucose uptake and serum insulin concentrations in diabetes patients.73 LY2405319, an FGF-21 agonist, highly improves lipid content in blood in T2D subjects with direct effect on controlling BMI, circulatory insulin levels, and production of adiponectin. FGF21 regulates metabolic balance by enhancing PPAR-γ actions by inducing the formation of beige or brown adipose tissue from white adipose tissue (WAT).74 Browning of WAT is a crucial balance involving two important factors-FGF-21 and irisin. Irisin is another prominent skeletal muscle-derived protein that has a significant role in MetS. Nutrition status has a direct effect on FGF-21 level in MetS. It suppresses the production and function of adiponectin and promotes the secretion of leptin and cytokine IL-6 in maturing adipocytes.75,76 High-sensitivity C-reactive protein (hsCRP, a popular indicator of inflammatory diseases) has intricate connection with FGF-21 in T2D subjects. Also, transcription and secretion of FGF-21 can be decreased by TNF-α. Circulatory FGF-21 concentrations are also linked to hypertension, making a role of FGF-21 in MetS quite significant. While pre-clinical studies were highly promising in terms of improving insulin sensitivity, the clinical development of FGF-21 as a drug was given up, as human studies showed significant FGF-21 resistance in T2D, undermining its efficacy in clinical T2D.77
4. Angiopoietin-related growth factor (AGF)
Angiopoietin-related growth factor (AGF) is produced from the liver and has crucial function in maintaining lipids and carbohydrates in our body. AGF deficiency is directly linked to, adiposity, T2D and fat depot formation in various organs. AGF also decreases hepatic gluconeogenesis. Serum AGF level increases in T2D patients and also in MetS patients. AGF increases with progression of T2D, while the level of HDLc in circulation decreases.78
5. Selenoprotein-P (SeP)
Selenoprotein-P is another crucial hepatokine. High level of SeP is found in T2D or fatty liver patients, or patients having visceral adipocyte accumulation. Such excess concentration of SeP is significantly correlated with IR, hsCRP, and triglyceride level. However, SeP in circulation is low in MetS subjects, particularly those with heart ailments.79,80 Moreover, excess concentrations of SeP decreases the risk of MetS in children. Adiponectin and salsalate inhibit SeP and this inhibition is mediated by AMPK-Forkhead box protein O1a (FOXO1 a) and its downstream proteins, thereby reducing hepatic glucose output.81 Serum SeP level shows inverse relation to serum adiponectin concentrations in T2D subjects. SeP can be a potent target for the management of IR-associated diseases.82
6. Angiopoietin-like proteins (ANGPTL)
Angiopoietin-like proteins (ANGPTL 1-8), are a group of proteins, secreted from the liver. These have a crucial role in maintaining lipid content in the body. First, we shall discuss about angiopoietin-like protein 4 (ANGPTL4). ANGPTL4 is also secreted from muscle cells and adipocytes. ANGPTL4 primarily regulates the storage of lipid in cells and its spilling. Because the lipid content is directly dependent on food intake, ANGPTL4 secretion is controlled by several factors such as the amount and time of food intake, nutrients present in food, and lifestyle.83 Nutrients in food, particularly fatty acids, can control gene expression of ANGPTL4 via PPARs. ANGPTL4 stimulates lipid breakdown within adipocytes through cAMP. ANGPTL4 is directly associated with IR. Excess production of ANGPTL4 is required to regulate insulin sensitivity; however, sometimes it may lead to excess lipid spillage in circulation and deposition of fat in various organs. ANGPTL4 concentration in circulation decreases in T2D patients, which implies that low level of ANGPTL4 can be a reason behind T2D. AGPTL4 has a key role in maintaining ROS level and inflammation.84 ANGPTL8 (also known as lipasin) is another newly studied liver-derived protein that regulates lipid metabolism. ANGPTL8 and HDL-cholesterol have a negative association, while ANGPTL8 and triglyceride level has a positive association. All the effects of ANGPTL8 are moderated by blocking the action of lipoprotein lipase and also ANGPTL3 by increasing the production of very low-density lipoprotein from hepatocytes. The expression of ANGPTL8 is directly related to an increased expression of hsCRP which in turn increases the prevalence of MetS. Plasma ANGPTL8 begin to increase in pre-diabetic patients and continue to increase until the progression of T2D.85 ANGPTL8 is also considered to be an indicator of kidney and insulin resistance. Increased secretion of ANGPTL8 occurs in MetS patients with hypertension along with increased expression of ANGPTL4.86 So, for future investigations ANGPTLs can be targeted by pharmaceuticals to treat hyperlipidemia.
7. Leukocyte cell-derived chemotaxin 2 (LECT2)
Leukocyte cell-derived chemotaxin 2 (LECT2), another hepatokine, is a key protein that can recognize energy demand in our body. It is connected to inflammation in the liver and natural killer T cells. LECT2 is also linked to fat accumulation and glucose transport impairment in skeletal muscles. Positive correlation exists between circulatory LECT2 concentrations, body weight, T2D, and increasing fat depot in the liver and other organs. Calorie-rich diet or fat-containing diet and workout or calorie expenditure exert opposite effects on LECT2 levels, increasing and decreasing its level, respectively, via blocking or activation the AMPK pathway.87 LECT2 treatment can increase inflammation by increasing the secretion of cytokines and other inflammatory molecules. LECT2 also increases insulin resistance, accumulation of fat, and phosphorylation of mammalian target of rapamycine (mTOR) in hepatocytes, making its role in MetS prominent.88
Conclusion
Taken altogether, adipokines and hepatokines likely contribute to insulin resistance, adiposity, T2D, cardiovascular diseases, dyslipidemia, hypertension, and inflammation, leading to MetS and its sinister consequences. Most adipokines and hepatokines are novel proteins. Several adipokines and hepatokines have crosstalk not only amongst themselves but also with other myokines or organokines. Better knowledge of all these proteins is the need of the hour to help us improve the preventive and therapeutic strategies to be adopted for MetS. The easiest and most effective way to restrain or suppress MetS is by making changes in our mode of lifestyle. Living a healthy life with proper food habits and exercise can delay the onset of MetS or aid in curing MetS along with medications.
Acknowledgments
AM is thankful to DBT (Department of Biotechnology, Research Associate Programme), Govt. of India, for her research fellowship (Award Number-DBT-RA/2022/January/N/1259).
Conflict of Interest
Dr. Satinath Mukhopadhyay reported Grants or contracts from AM is thankful to Science & Engineering Research Board (SERB), Department of Science & Technology, Govt. of India, for her research fellowship (Grant No. ECR/2017/001028/LS). SM and SAM are thankful to the Department of Science and Technology, Government of West Bengal, Bikash Bhawan, Bidhan Nagar, Kolkata, India [853(Sanc.)/ST/P/S&T/9G-2/2011] for funding.
All other authors reported no conflict of Interest.
References
- Metabolic syndrome pandemic. Arterioscler Thromb Vasc Biol. 2008;28(04):629-636. DOI: 10.1161/ATVBAHA.107.151092
- [CrossRef] [PubMed] [Google Scholar]
- Insulin resistance. A multifaceted syndrome responsible for NIDDM, obesity, hypertension, dyslipidemia, and atherosclerotic cardiovascular disease. Diabetes Care. 1991;14(03):173-194. DOI: 10.2337/diacare.14.3.173
- [CrossRef] [PubMed] [Google Scholar]
- Definition, diagnosis and classification of diabetes mellitus and its complications : report of a WHO consultation. Part 1, Diagnosis and classification of diabetes mellitus Accessed November 27, 2011, at: https://apps.who.int/iris/handle/10665/66040
- [Google Scholar]
- European Group for the Study of Insulin Resistance (EGIR). Comment on the provisional report from the WHO consultation. Diabet Med. 1999;16(05):442-443. DOI: 10.1046/j.1464-5491.1999.00059.x
- [CrossRef] [PubMed] [Google Scholar]
- Executive summary of the third report of The National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, And Treatment of High Blood Cholesterol In Adults (Adult Treatment Panel III) JAMA. 2001;285(19):2486-2497. DOI: 10.1001/jama.285.19.2486
- [CrossRef] [PubMed] [Google Scholar]
- Prevalence and risk factors associated with the metabolic syndrome and dyslipidemia in White, Black, Amerindian and Mixed Hispanics in Zulia State, Venezuela. Diabetes Res Clin Pract. 2005;69(01):63-77. DOI: 10.1016/j.diabres.2004.11.018
- [CrossRef] [PubMed] [Google Scholar]
- Metabolic syndrome determinants in an urban population from Brazil: social class and gender-specific interaction. Int J Cardiol. 2008;129(02):259-265. DOI: 10.1016/j.ijcard.2007.07.097
- [CrossRef] [PubMed] [Google Scholar]
- Adiponectin secretion and response to pioglitazone is depot dependent in cultured human adipose tissue. Am J Physiol Endocrinol Metab. 2008;295(04):E842-E850. DOI: 10.1152/ajpendo.90359.2008
- [CrossRef] [PubMed] [Google Scholar]
- Insulin resistance, hyperinsulinemia, hypertriglyceridemia, and hypertension. Parallels between human disease and rodent models. Diabetes Care. 1991;14(03):195LP-202. DOI: 10.2337/diacare.14.3.195
- [CrossRef] [PubMed] [Google Scholar]
- Metabolic syndrome and incident diabetes: current state of the evidence. Diabetes Care. 2008;31(09):1898-1904. DOI: 10.2337/dc08-0423
- [CrossRef] [PubMed] [Google Scholar]
- Triglycerides are major determinants of cholesterol esterification/transfer and HDL remodeling in human plasma. Arterioscler Thromb Vasc Biol. 1995;15(11):1819-1828. DOI: 10.1161/01.ATV.15.11.1819
- [CrossRef] [PubMed] [Google Scholar]
- Worldwide prevalence of hypertension: a systematic review. J Hypertens. 2004;22(01):11-19. DOI: 10.1097/00004872-200401000-00003
- [CrossRef] [PubMed] [Google Scholar]
- Hyperresistinemia is associated with coexistence of hypertension and type 2 diabetes. Hypertens (Dallas, Tex 1979). 2008;51(02):534-539. DOI: 10.1161/HYPERTENSIONAHA.107.103077
- [CrossRef] [PubMed] [Google Scholar]
- Epidemiology and prevention of hypertension. Med Clin North Am. 1997;81(05):1077-1097. DOI: 10.1016/s0025-7125(05)70568-x
- [CrossRef] [PubMed] [Google Scholar]
- Leptin-replacement therapy for lipodystrophy. N Engl J Med. 2002;346(08):570-578. DOI: 10.1056/NEJMoa012437
- [CrossRef] [PubMed] [Google Scholar]
- Effects of recombinant leptin therapy in a child with congenital leptin deficiency. N Engl J Med. 1999;341(12):879-884. DOI: 10.1056/NEJM199909163411204
- [CrossRef] [PubMed] [Google Scholar]
- Dipeptidyl peptidase 4 is a novel adipokine potentially linking obesity to the metabolic syndrome. Diabetes. 2011;60(07):1917-1925. DOI: 10.2337/db10-1707
- [CrossRef] [PubMed] [Google Scholar]
- Adipose dipeptidyl peptidase-4 and obesity: correlation with insulin resistance and depot-specific release from adipose tissue in vivo and in vitro. Diabetes Care. 2013;36(12):4083-4090. DOI: 10.2337/dc13-0496
- [CrossRef] [PubMed] [Google Scholar]
- Adipokines: potential therapeutic targets for vascular dysfunction in type II diabetes mellitus and obesity. J Diabetes Res. 2017;2017:8095926. DOI: 10.1155/2017/8095926
- [CrossRef] [PubMed] [Google Scholar]
- Adipose tissue resistin expression is severely suppressed in obesity and stimulated by peroxisome proliferator-activated receptor gamma agonists. J Biol Chem. 2001;276(28):25651-25653. DOI: 10.1074/jbc.C100189200
- [CrossRef] [PubMed] [Google Scholar]
- Resistin and obesity-associated insulin resistance. Trends Endocrinol Metab. 2002;13(01):18-23. DOI: 10.1016/s1043-2760(01)00522-7
- [CrossRef] [PubMed] [Google Scholar]
- Serum resistin (FIZZ3) protein is increased in obese humans. J Clin Endocrinol Metab. 2004;89(04):1977. author reply 1977–1978 DOI: 10.1210/jc.2003-031988
- [CrossRef] [PubMed] [Google Scholar]
- Adenovirus-mediated chronic “hyper-resistinemia” leads to in vivo insulin resistance in normal rats. J Clin Invest. 2004;114(02):224-231. DOI: 10.1172/JCI20785
- [CrossRef] [PubMed] [Google Scholar]
- Roles of adipocyte derived hormone adiponectin and resistin in insulin resistance of type 2 diabetes. World J Gastroenterol. 2006;12(11):1747-1751. DOI: 10.3748/wjg.v12.i11.1747
- [CrossRef] [PubMed] [Google Scholar]
- Oligomeric resistin impairs insulin and AICAR-stimulated glucose uptake in mouse skeletal muscle by inhibiting GLUT4 translocation. Am J Physiol Endocrinol Metab. 2009;297(01):E57-E66. DOI: 10.1152/ajpendo.90744.2008
- [CrossRef] [PubMed] [Google Scholar]
- Association between serum resistin concentration and hypertension: A systematic review and meta-analysis. Oncotarget. 2017;8(25):41529-41537. DOI: 10.18632/oncotarget.17561
- [CrossRef] [PubMed] [Google Scholar]
- Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-alpha, in vivo. J Clin Endocrinol Metab. 1997;82(12):4196-4200. DOI: 10.1210/jcem.82.12.4450
- [CrossRef] [Google Scholar]
- Point-counterpoint: interleukin-6 does/does not have a beneficial role in insulin sensitivity and glucose homeostasis. J Appl Physiol. 2007;102(02):820-821. author reply 825 DOI: 10.1152/japplphysiol.01353.2006
- [CrossRef] [Google Scholar]
- A stress signaling pathway in adipose tissue regulates hepatic insulin resistance. Science. 2008;322(5907):1539-1543. DOI: 10.1126/science.1160794
- [CrossRef] [PubMed] [Google Scholar]
- Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes. 2007;56(04):1010-1013. DOI: 10.2337/db06-1656
- [Google Scholar]
- Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. Am J Physiol Endocrinol Metab. 2001;280(05):E745-E751. DOI: 10.1152/ajpendo.2001.280.5.E745
- [CrossRef] [PubMed] [Google Scholar]
- Signaling specificity of interleukin-6 action on glucose and lipid metabolism in skeletal muscle. Mol Endocrinol. 2006;20(12):3364-3375. DOI: 10.1210/me.2005-0490
- [CrossRef] [PubMed] [Google Scholar]
- Impact of intensive lifestyle modification on levels of adipokines and inflammatory biomarkers in metabolically healthy obese women. Mediators Inflamm. 2019;2019:4165260. DOI: 10.1155/2019/4165260
- [CrossRef] [PubMed] [Google Scholar]
- Abdominal obesity and metabolic syndrome. Nature. 2006;444(7121):881-887. DOI: 10.1038/nature05488
- [CrossRef] [PubMed] [Google Scholar]
- Local and systemic insulin resistance resulting from hepatic activation of IKK-beta and NF-kappaB. Nat Med. 2005;11(02):183-190. DOI: 10.1038/nm1166
- [CrossRef] [PubMed] [Google Scholar]
- Protection from obesity-induced insulin resistance in mice lacking TNF-alpha function. Nature. 1997;389(6651):610-614. DOI: 10.1038/39335
- [CrossRef] [PubMed] [Google Scholar]
- Tumor necrosis factor alpha: a key component of the obesity-diabetes link. Diabetes. 1994;43(11):1271-1278. DOI: 10.2337/diab.43.11.1271
- [CrossRef] [PubMed] [Google Scholar]
- TNF-alpha induction of lipolysis is mediated through activation of the extracellular signal related kinase pathway in 3T3-L1 adipocytes. J Cell Biochem. 2003;89(06):1077-1086. DOI: 10.1002/jcb.10565
- [CrossRef] [PubMed] [Google Scholar]
- TNF-α/IL-10 ratio: An independent predictor for coronary artery disease in North Indian population. Diabetes Metab Syndr. 2018;12(03):221-225. DOI: 10.1016/j.dsx.2017.09.006
- [CrossRef] [PubMed] [Google Scholar]
- Genetic variation in the gene encoding adiponectin is associated with an increased risk of type 2 diabetes in the Japanese population. Diabetes. 2002;51(02):536-540. DOI: 10.2337/diabetes.51.2.536
- [CrossRef] [PubMed] [Google Scholar]
- The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med. 2001;7(08):941-946. DOI: 10.1038/90984
- [CrossRef] [PubMed] [Google Scholar]
- Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med. 2002;8(11):1288-1295. DOI: 10.1038/nm788
- [CrossRef] [PubMed] [Google Scholar]
- Total and high-molecular-weight adiponectin and resistin in relation to the risk for type 2 diabetes in women. Ann Intern Med. 2008;149(05):307-316. DOI: 10.7326/0003-4819-149-5-200809020-00005
- [CrossRef] [PubMed] [Google Scholar]
- Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation. 2001;103(08):1057-1063. DOI: 10.1161/01.cir.103.8.1057
- [CrossRef] [PubMed] [Google Scholar]
- AdipoQ is a novel adipose-specific gene dysregulated in obesity. J Biol Chem. 1996;271(18):10697-10703. DOI: 10.1074/jbc.271.18.10697
- [CrossRef] [PubMed] [Google Scholar]
- Adiponectin in youth: relationship to visceral adiposity, insulin sensitivity, and beta-cell function. Diabetes Care. 2004;27(02):547-552. DOI: 10.2337/diacare.27.2.547
- [CrossRef] [PubMed] [Google Scholar]
- Kaempferol alleviates palmitic acid-induced lipid stores, endoplasmic reticulum stress and pancreatic β-cell dysfunction through AMPK/mTOR-mediated lipophagy. J Nutr Biochem. 2018;57:212-227. DOI: 10.1016/j.jnutbio.2018.02.017
- [CrossRef] [PubMed] [Google Scholar]
- Effect of a weight loss program on serum adiponectin and insulin resistance among overweight and obese premenopausal females. J Egypt Public Health Assoc. 2020;95(01):32. DOI: 10.1186/s42506-020-00060-z
- [CrossRef] [PubMed] [Google Scholar]
- Weight-loss diets, adiponectin, and changes in cardiometabolic risk in the 2-Year POUNDS lost trial. J Clin Endocrinol Metab. 2016;101(06):2415-2422. DOI: 10.1210/jc.2016-1207
- [CrossRef] [PubMed] [Google Scholar]
- Effect of diet on adiponectin levels in blood. Nutr Rev. 2011;69(10):599-612. DOI: 10.1111/j.1753-4887.2011.00414.x
- [CrossRef] [PubMed] [Google Scholar]
- Addition of exercise increases plasma adiponectin and release from adipose tissue. Med Sci Sports Exerc. 2015;47(11):2450-2455. DOI: 10.1249/MSS.0000000000000670
- [CrossRef] [PubMed] [Google Scholar]
- The discovery and development of liraglutide and semaglutide. Front Endocrinol (Lausanne). 2019;10:155. DOI: 10.3389/fendo.2019.00155
- [CrossRef] [PubMed] [Google Scholar]
- Effects of novel dual GIP and GLP-1 receptor agonist tirzepatide on biomarkers of nonalcoholic steatohepatitis in patients with type 2 diabetes. Diabetes Care. 2020;43(06):1352-1355. DOI: 10.2337/dc19-1892
- [CrossRef] [PubMed] [Google Scholar]
- Direct inhibitory effects of pioglitazone on hepatic fetuin-A expression. PLoS One. 2014;9(02):e88704. DOI: 10.1371/journal.pone.0088704
- [CrossRef] [PubMed] [Google Scholar]
- Rosiglitazone and pioglitazone similarly improve insulin sensitivity and secretion, glucose tolerance and adipocytokines in type 2 diabetic patients. Diabetes Obes Metab. 2008;10(12):1204-1211. DOI: 10.1111/j.1463-1326.2008.00880.x
- [CrossRef] [PubMed] [Google Scholar]
- Adipose tissue as an endocrine organ: impact on insulin resistance. Neth J Med. 2003;61(06):194-212.
- [Google Scholar]
- Molecular and anatomical determinants of central leptin resistance. Nat Neurosci. 2005;8(05):566-570. DOI: 10.1038/nn1454
- [CrossRef] [PubMed] [Google Scholar]
- Diet- and diabetes-induced changes of ob gene expression in rat adipose tissue. FEBS Lett. 1995;371(03):324-328. DOI: 10.1016/0014-5793(95)00943-4
- [CrossRef] [PubMed] [Google Scholar]
- Recent advances in understanding leptin signaling and leptin resistance. Am J Physiol Endocrinol Metab. 2009;297(06):E1247-E1259. DOI: 10.1152/ajpendo.00274.2009
- [CrossRef] [PubMed] [Google Scholar]
- Non-responders after dastric bypass surgery for morbid obesity: peptide hormones and glucose homeostasis. Obes Surg. 2019;29(12):4008-4017. DOI: 10.1007/s11695-019-04089-8
- [CrossRef] [PubMed] [Google Scholar]
- Leptin and obesity: role and clinical implication. Front Endocrinol (Lausanne). 2021;12:585887. DOI: 10.3389/fendo.2021.585887
- [CrossRef] [PubMed] [Google Scholar]
- Reversal of type 2 diabetes: normalisation of beta cell function in association with decreased pancreas and liver triacylglycerol. Diabetologia. 2011;54(10):2506-2514. DOI: 10.1007/s00125-011-2204-7
- [CrossRef] [PubMed] [Google Scholar]
- Primary care-led weight management for remission of type 2 diabetes (DiRECT): an open-label, cluster-randomised trial. Lancet. 2018;391(10120):541-551. DOI: 10.1016/S0140-6736(17)33102-1
- [CrossRef] [PubMed] [Google Scholar]
- Leptin stimulates fatty-acid oxidation by activating AMP-activated protein kinase. Nature. 2002;415(6869):339-343. DOI: 10.1038/415339a
- [CrossRef] [PubMed] [Google Scholar]
- Role of obesity and leptin in the pubertal process and pubertal growth–a review. Int J Obes Relat Metab Disord. 2003;27(08):869-874. DOI: 10.1038/sj.ijo.0802328
- [CrossRef] [PubMed] [Google Scholar]
- Hepatokines: unlocking the multi-organ network in metabolic diseases. Diabetologia. 2015;58(08):1699-1703. DOI: 10.1007/s00125-015-3634-4
- [CrossRef] [PubMed] [Google Scholar]
- Liver-targeting drugs and their effect on blood glucose and hepatic lipids. Diabetologia. 2021;64(07):1461-1479. DOI: 10.1007/s00125-021-05442-2
- [CrossRef] [PubMed] [Google Scholar]
- Metabolic adaptation in obesity and type II diabetes: myokines, adipokines and hepatokines. Int J Mol Sci. 2016;18(01):E8. DOI: 10.3390/ijms18010008
- [CrossRef] [PubMed] [Google Scholar]
- Implication of hepatokines in metabolic disorders and cardiovascular diseases. BBA Clin. 2016;5:108-113. DOI: 10.1016/j.bbacli.2016.03.002
- [CrossRef] [PubMed] [Google Scholar]
- The effects of caloric restriction on fetuin-A and cardiovascular risk factors in rats and humans: a randomized controlled trial. Clin Endocrinol (Oxf). 2013;79(03):356-363. DOI: 10.1111/cen.12076
- [CrossRef] [PubMed] [Google Scholar]
- Exercise-induced lowering of fetuin-A may increase hepatic insulin sensitivity. Med Sci Sports Exerc. 2014;46(11):2085-2090. DOI: 10.1249/MSS.0000000000000338
- [CrossRef] [PubMed] [Google Scholar]
- A novel hepatokine, HFREP1, plays a crucial role in the development of insulin resistance and type 2 diabetes. Diabetologia. 2016;59(08):1732-1742. DOI: 10.1007/s00125-016-3991-7
- [CrossRef] [PubMed] [Google Scholar]
- Direct effects of FGF21 on glucose uptake in human skeletal muscle: implications for type 2 diabetes and obesity. Diabetes Metab Res Rev. 2011;27(03):286-297. DOI: 10.1002/dmrr.1177
- [CrossRef] [PubMed] [Google Scholar]
- FGF21 regulates PGC-1α and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012;26(03):271-281. DOI: 10.1101/gad.177857.111
- [CrossRef] [PubMed] [Google Scholar]
- Fibroblast growth factor 21 is elevated in metabolically unhealthy obesity and affects lipid deposition, adipogenesis, and adipokine secretion of human abdominal subcutaneous adipocytes. Mol Metab. 2015;4(07):519-527. DOI: 10.1016/j.molmet.2015.04.002
- [CrossRef] [PubMed] [Google Scholar]
- Inflammatory markers in obese adolescents with type 2 diabetes and their relationship to hepatokines and adipokines. J Pediatr. 2016;173:131-135. DOI: 10.1016/j.jpeds.2016.02.055
- [CrossRef] [PubMed] [Google Scholar]
- Elevated serum fibroblast growth factor 21 is associated with hypertension in community-dwelling adults. J Hum Hypertens. 2013;27(06):397-399. DOI: 10.1038/jhh.2012.52
- [CrossRef] [PubMed] [Google Scholar]
- Relationship between serum levels of angiopoietin-related growth factor and metabolic risk factors. Horm Metab Res = Horm und Stoffwechselforsch = Horm Metab. 2014;46(10):685-690. DOI: 10.1055/s-0034-1382078
- [CrossRef] [PubMed] [Google Scholar]
- Serum selenoprotein P levels in patients with type 2 diabetes and prediabetes: implications for insulin resistance, inflammation, and atherosclerosis. J Clin Endocrinol Metab. 2011;96(08):E1325-E1329. DOI: 10.1210/jc.2011-0620
- [CrossRef] [PubMed] [Google Scholar]
- Association of expression of selenoprotein P in mRNA and protein levels with metabolic syndrome in subjects with cardiovascular disease: results of the Selenegene study. J Gene Med. 2017;19(03) DOI: 10.1002/jgm.2945
- [CrossRef] [PubMed] [Google Scholar]
- Salsalate and adiponectin improve palmitate-induced insulin resistance via inhibition of selenoprotein P through the AMPK-FOXO1α pathway. PLoS One. 2013;8(06):e66529. DOI: 10.1371/journal.pone.0066529
- [CrossRef] [PubMed] [Google Scholar]
- Inverse correlation between serum levels of selenoprotein P and adiponectin in patients with type 2 diabetes. PLoS One. 2012;7(04):e34952. DOI: 10.1371/journal.pone.0034952
- [CrossRef] [PubMed] [Google Scholar]
- The fasting-induced adipose factor/angiopoietin-like protein 4 is physically associated with lipoproteins and governs plasma lipid levels and adiposity. J Biol Chem. 2006;281(02):934-944. DOI: 10.1074/jbc.M506519200
- [CrossRef] [PubMed] [Google Scholar]
- Angiopoietin-like protein 4 decreases blood glucose and improves glucose tolerance but induces hyperlipidemia and hepatic steatosis in mice. Proc Natl Acad Sci U S A. 2005;102(17):6086-6091. DOI: 10.1073/pnas.0408452102
- [CrossRef] [PubMed] [Google Scholar]
- Increased serum ANGPTL8 concentrations in patients with prediabetes and Type 2 diabetes. J Diabetes Res. 2017;2017:8293207. DOI: 10.1155/2017/8293207
- [CrossRef] [PubMed] [Google Scholar]
- Increased plasma and adipose tissue levels of ANGPTL8/betatrophin and ANGPTL4 in people with hypertension. Lipids Health Dis. 2018;17(01):35. DOI: 10.1186/s12944-018-0681-0
- [CrossRef] [PubMed] [Google Scholar]
- LECT2 functions as a hepatokine that links obesity to skeletal muscle insulin resistance. Diabetes. 2014;63(05):1649LP-1664. DOI: 10.2337/db13-0728
- [CrossRef] [PubMed] [Google Scholar]
- A dipeptidyl peptidase-IV inhibitor improves hepatic steatosis and insulin resistance by AMPK-dependent and JNK-dependent inhibition of LECT2 expression. Biochem Pharmacol. 2015;98(01):157-166. DOI: 10.1016/j.bcp.2015.08.098
- [CrossRef] [PubMed] [Google Scholar]



