
Translate this page into:
A Novel Antigonadotropic Role of Thyroid Stimulating Hormone on Leydig Cell-Derived Mouse Leydig Tumor Cells-1 Line
Address for correspondence Skand Shekhar, MD, Section on Endocrinology and Genetics, National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, MD 20892, United States (e-mail: skand.shekhar@nih.gov).
This article was originally published by Thieme Medical and Scientific Publishers Pvt. Ltd. and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Subclinical hypothyroid men characterized by a rise in only thyroid stimulating hormone (TSH) levels, and normal thyroid hormone levels showed a fall in their serum progesterone and testosterone levels. This suggested a role of TSH in regulating Leydig cell steroidogenesis. Therefore, we investigated the direct role of TSH on steroid production and secretion using a mouse Leydig tumor cell line-1 (MLTC-1). MLTC-1 cells were treated with different doses of TSH isolated from porcine pituitary as well as recombinant TSH. Steroid secretion was measured by radioimmunoassay (RIA). The mRNA levels of steroidogenic enzymes were quantitated by real-time polymerase chain reaction (RT-PCR), whereas the corresponding protein levels were determined by western blot. In MLTC-1 cells, pituitary TSH as well as recombinant TSH inhibited progesterone and testosterone secretion in a dose-dependent manner. The inhibitory action of TSH on steroid secretion was unique and not mimicked by other anterior pituitary hormones including follicle stimulating hormone and adrenocorticotropic hormone. Recombinant TSH showed no effect on steroidogenic acute regulatory protein and CYP11A1, the enzymes catalyzing the nonsteroidogenic and steroidogenic rate-limiting steps of steroid synthesis, respectively. Recombinant TSH was shown to inhibit steroidogenesis in MLTC-1 cells by inhibiting the 3-β hydroxy steroid dehydrogenase mRNA and protein levels, the enzyme that catalyzes the conversion of pregnenolone to progesterone. This inhibitory effect of TSH is probably direct as both mRNA and protein of the TSH receptor were shown to be present in the MLTC-1 cells.
Keywords
Leydig cells
steroidogenesisthyroid stimulating hormone receptor
thyroid stimulating hormone

Introduction
The thyroid hormone 3, 5, 3′-L-triiodothyronine (T3) was shown to increase the basal and 3′, 5′ cyclic adenosine monophosphate (cAMP)-stimulated steroid production in primary Leydig cells and their derived lines.1-3 Overt hypothyroid men showed a significant reduction in their serum progesterone and testosterone levels.4 This reduction in serum steroid levels could be due to reduced thyroid hormone levels or increased thyroid stimulating hormone (TSH) levels or both. Although animal studies have consistently showed the direct effects of T3 on Leydig cell function in the form of cell proliferation and steroidogenesis; however, the exact effects and mechanisms of action remain controversial.5,6 Inconsistencies in the findings of existing literature about the impact of hypothyroidism on testosterone and gonadotropin production raise questions about the role of TSH in vivo.7,8 Studies in subclinical hypothyroid men, characterized by a rise in only TSH levels with normal thyroid hormone levels, also showed a similar reduction in serum progesterone and testosterone levels.9 As only TSH was raised in these subclinical hypothyroid men, this furthers the suggestion that a direct inhibitory action of TSH occurs on steroid production. Interestingly, studies have demonstrated the presence of TRH receptors on Leydig cells, which is unique to them among all testicular cell types.10 This also raises the possibility of other thyroid function modulating hormones such as TSH, acting on Leydig cells through their receptors. To date, no studies have been conducted to see the direct modulation of steroid production and secretion by TSH in Leydig cells.
Therefore, we investigated the role of TSH on steroid production and secretion using a mouse Leydig tumor cell line-1 (MLTC-1). We also investigated the effect of TSH on steroidogenic enzymes to dissect out the molecular mechanism(s) used by TSH for regulating steroid secretion.
Materials and Methods
Ethics Clearance
The study protocol was approved by the Institute Ethics committee of All India Institute of Medical Sciences.
Hormones and Chemicals
Pituitary-derived TSH, luteinizing hormone (LH), follicle-stimulating hormone (FSH), and adrenocorticotropic hormone (ACTH) were purchased from Sigma–Aldrich (St. Louis, Missouri, United States). Recombinant TSH was purchased from R&D Systems (Minneapolis, Minnesota, United States). Tritium-labeled radioactive progesterone and testosterone were purchased from Perkin Elmer (Waltham, Massachusetts, United States). Both the mouse monoclonal testosterone antibody (clone 4E1G2) and rabbit polyclonal progesterone antibody were purchased from Bio-Rad (Hercules, California, United States). Primers were purchased from Sigma–Aldrich. Rabbit antisteroidogenic acute regulatory protein (StAR) antibody and rabbit anti-CYP11A1 antibody were purchased from Cell Signaling Technologies (Boston, Massachusetts, United States). Rabbit anti-3β-hydroxy steroid dehydrogenase-1 (3β-HSD) antibody and rabbit anti-TSH receptor antibody were purchased from Abcam (Cambridge, United Kingdom). Antirabbit HRP-labeled secondary antibody was purchased from Cell Signaling Technologies.
Cell Culture
MLTC-1 cells were obtained from the American Type Culture Collection (Manassas, Virginia, United States). MLTC-1 cells express the human chorionic gonadotropin (hCG) receptor. They respond to LH/hCG with stimulation of steroid production.11 The MLTC-1 cells were routinely maintained in Waymouth MB 752/1 medium (Sigma–Aldrich) supplemented with 10% heat-inactivated fetal bovine serum (Sigma–Aldrich, St. Louis, Missouri, United States) at 37°C in 5% CO2 in a humidified incubator. The MLTC-1 cells used for all the experiments had a passage number less than 30.
Cell Treatments
MLTC-1 cells were seeded at a density of 2 × 105 cells/mL/well for measuring steroid secretion and at 1 × 106 cells/well for total RNA extraction and western blot assays. MLTC-1 cells were preincubated separately for 2 hours with different doses of pituitary or recombinant TSH, or FSH or ACTH in serum free media. Thereafter, the cells were incubated with 100 mIU/mL LH or 1 mM 8-Br-cAMP. The total period of incubation including the time of preincubation was 6 hours as MLTC-1 cells begin proliferation by 9 hours. Viability of the cells after treatment were tested using [3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide] (MTT, Sigma–Aldrich, St. Louis, Missouri, United States).
Steroid Production
Steroid secretion was measured in cell supernatant by RIA according to the World Health Organization guidelines. Both the mouse monoclonal testosterone antibody (clone 4E1G2) and rabbit polyclonal progesterone antibody were used at a dilution of 1:500. The progesterone antibody used for RIA was highly specific and showed no crossreactivity for pregnenolone as well as for testosterone. However, the testosterone antibody also recognizes 5α-dihydrotestosterone (DHT, 1%) and androstenedione (1%). The intraassay variation was less than 5%, and the interassay variation was less than 10%.
RNA Extraction and Reverse Transcriptase Polymerase Chain Reaction
Total RNA was extracted using Ribozol RNA extraction reagent (Amresco, Solon, Ohio, United States) as per manufacturer's instructions. Total RNA isolated from the cells was reverse transcribed using oligo dT primers and Revert Aid First Strand cDNA Synthesis Kit (Thermo Scientific, Waltham, Massachusetts, United States). The cDNA synthesized was amplified using gene-specific primers (Sigma–Aldrich, St. Louis, United States). The sequences of StAR primers were forward 5′ GACCTTGAAAGGCTCAGGAAGAAC 3′ and reverse 5′ TAGCTGAAGATGGACAGACTTGC 3′.1 The sequences of CYP11A1 primers were forward 5′ CAACATCACAGAGATGCTGGCAGG3′ and reverse 5′ CTCAGGCATCAGGATGAGGTTGAA 3′.12 The sequences of 3β-HSD-I primers were forward 5′ GACAGGAGCAGGAGGGTTTGTGG 3′ and reverse 5′ CTCCTTCTAACATTGTCACCTTGGCCT 3′.13 The sequences of TSH receptor (TSHR) primers were forward 5′ TTC CAG CCG CTG CAG AGT TGC3′ and reverse 5′ GAG TGT GCG TCT CCA CCC TG 3′.14 The sequences of GAPDH primers were forward 5′ ACGGGAAGCTTGTCATCAAT 3′ and reverse 5′TGGACTCCACGACGTCGTACTCA 3′.15 The steroidogenic enzymes were amplified by quantitative RT-PCR. The PCR conditions used were initial denaturation at 95°C for 5 minutes, which followed by 40 cycles of 95°C for 20 seconds, 56°C for 20 seconds, and 72°C for 40 seconds. Ct values obtained during the RT-PCR reaction were used to calculate the relative mRNA expression using the formula: relative mRNA expressionStAR/CYP11A1/3β-HSD = 2−ΔΔCt, where ΔCt = CtStAR/CYP11A1/3β-HSD–CtGAPDH and ΔΔCt = ΔCtBasal–ΔCtTreated. The TSHR gene was amplified by semiquantitative RT-PCR. The PCR conditions used were initial denaturation at 95°C for 3 minutes, which followed by 40 cycles of 95°C for 30 seconds, 60°C for 20 seconds, and 72°C for 45 seconds. The amplified products were size-fractionated in a 2% (w/v) agarose gel and visualized by staining with ethidium bromide (0.2 ng/mL).
Western Blot
Total protein extracts were separated by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS/PAGE) using 12% resolving gel. The protein bands in the gel were transferred onto a 0.45-µm polyvinylidene fluoride membrane. Polyvinylidene fluoride membrane was blocked with 5% bovine serum albumin solution made in TBS-T (0.1% Tween-20 in 1× TBS). The membrane was probed with rabbit anti-StAR antibody, rabbit anti-CYP11A1 antibody, rabbit anti-3β- HSD antibody, rabbit anti-TSHR antibody, or rabbit anti-β-actin antibody. A 1:1,000 dilution was used for each of the primary antibody and incubated for overnight at 4°C. Antirabbit HRP-labeled secondary antibody at 1:3,000 dilution was used. Protein bands were visualized using ECL system (Merck Millipore, Billerica, United States). The density of the protein bands was measured using ImageJ software developed at the National Institute of Health (Bethesda, Maryland, United States).
Statistical Analysis
All the treatments were performed in triplicates and at least three times. The data were pooled and analyzed using GraphPad Prism 4.0 software (GraphPad Prism Software Inc., San Diego, California, United States). The results were analyzed by Student's t-test. A p-value <0.05 was considered significant.
Results
TSH Inhibits Steroid Secretion
Initially, different doses of pituitary TSH ranging from 1 to 100 mIU/mL were tested for their effect on steroid secretion by MLTC-1 cells. The minimum dose of 1 mIU/mL of TSH showed no significant effect on basal, LH or 8-Br-cAMP-stimulated progesterone and testosterone secretion. However, the higher doses of TSH significantly inhibited both progesterone and testosterone secretion in a dose dependent manner (►Fig. 1A, B). About 100 mIU/mL of TSH inhibited basal progesterone and testosterone secretion by 37 and 39%, respectively (►Fig. 1A, B). 100mIU/ml of TSH also inhibited LH and 8-Br-cAMP-stimulated progesterone secretion by 35 and 25%, respectively. About 100 mIU/mL of TSH inhibited LH and 8-Br-cAMP-stimulated testosterone secretion by 42 and 27%, respectively (►Fig. 1A, B).

- Effect of pituitary TSH on (A) progesterone (B) testosterone secretion by mouse Leydig tumor cell line-1 cells under basal and treated conditions. Luteinizing hormone or 8-Br-cAMP was added after 2-hour preincubation with different doses of TSH. Bars represent mean ± standard deviation of three experiments; each experiment was done at least in triplicates (n = 10). *indicate p < 0.05; **indicate p < 0.01; and ***indicate p < 0.001 in cells treated without and with TSH in the corresponding groups. TSH, thyroid stimulating hormone.
Effect of TSH on Steroid Secretion is Specific
To rule out the possibility that the inhibitory effect of TSH on steroid secretion is nonspecific in nature, we used two other pituitary-derived hormones: FSH and ACTH. However, both FSH and ACTH showed no significant effect on basal, LH or 8-Br-cAMP-stimulated progesterone and testosterone secretion (►Fig. 2A–D). This suggested that steroid secretion is specifically inhibited by TSH.
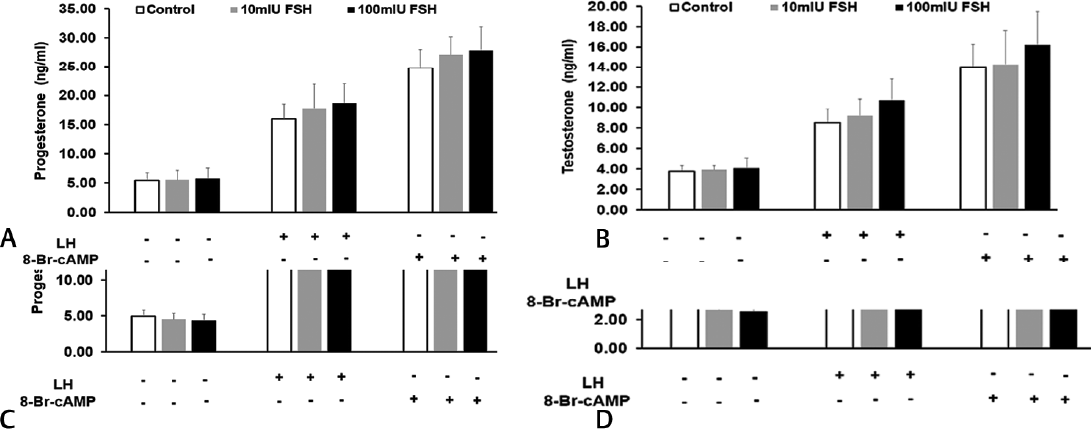
- Effect of pituitary FSH on (A) progesterone (B) testosterone secretion; effect of pituitary ACTH on (C) progesterone and (D) testosterone secretion by mouse Leydig tumor cell line-1 cells under basal and treated conditions. LH or 8-Br-cAMP were added after 2 hour preincubation with different doses of FSH or ACTH. Bars represent mean ± standard deviation of three experiments; each experiment was done at least in triplicates (n = 10). ACTH, adrenocorticotropic hormone; FSH, follicle-stimulating hormone.
To further confirm that the inhibitory effect of TSH was not due to any other contaminant present in the pituitary extract, we used recombinant TSH (rTSH). The minimum dose of 0.15 ng/mL of rTSH showed no significant effect on basal or 8-Br-cAMP-stimulated progesterone and testosterone secretion. However, both the higher doses of rTSH significantly inhibited both progesterone and testosterone secretion in a dose-dependent manner (►Fig. 3A, B). While 1.5 ng/mL of rTSH inhibited basal progesterone and testosterone secretion by 32 and 31%, 15 ng/mL of rTSH inhibited those by 61 and 45%, respectively (►Fig. 3A, B). The inhibitory effect of rTSH on 8-Br-cAMP-stimulated progesterone and testosterone secretion was even more pronounced. Maximal inhibition was observed with 15 ng/mL of rTSH on 8-Br-cAMP-stimulated progesterone and testosterone secretion by 48 and 49%, respectively (►Fig. 3A, B).
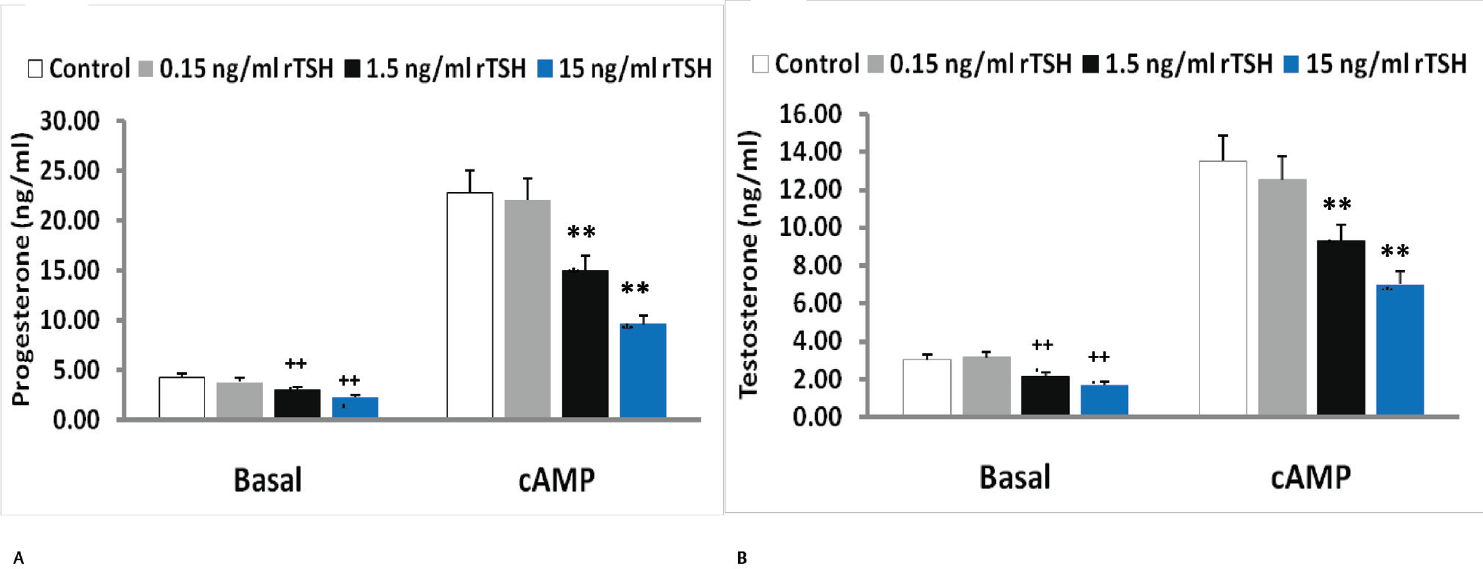
- Effect of recombinant TSH (rTSH) on (A) progesterone (B) testosterone secretion by mouse Leydig tumor cell line-1 cells under basal and treated conditions. 8-Br-cAMP was added after 2-hour preincubation with different doses of TSH. Bars represent mean ± standard deviation of three experiments; each experiment was done at least in triplicates (n = 10). +++indicate p < 0.001 in cells treated without and with TSH in the basal group. ***indicate p < 0.001 in cells treated without and with TSH in the 8-Br-cAMP treated group. TSH, thyroid stimulating hormone.
Effect of TSH on Expression of Steroidogenic Enzymes
The molecular mechanism underlying the inhibition of steroid secretion by TSH was worked out by studying the expression of the steroidogenic enzymes after treatment with rTSH. Interestingly, rTSH showed no significant effect on StAR and CYP11A1 mRNA and protein levels in MLTC-1 cells (►Figs. 4A, B and 5A, B), but expression of 3β-HSD was significantly reduced by both 1.5 and 15 ng/mL dose of rTSH (►Figs. 4C and 5C). Total 15 ng/mL of rTSH inhibited basal and 8-Br-cAMP-stimulated 3β-HSD1-I mRNA levels by 37 and 51%, respectively (►Fig. 4C), while the protein levels were inhibited by 40 and 53%, respectively (►Fig. 5C).
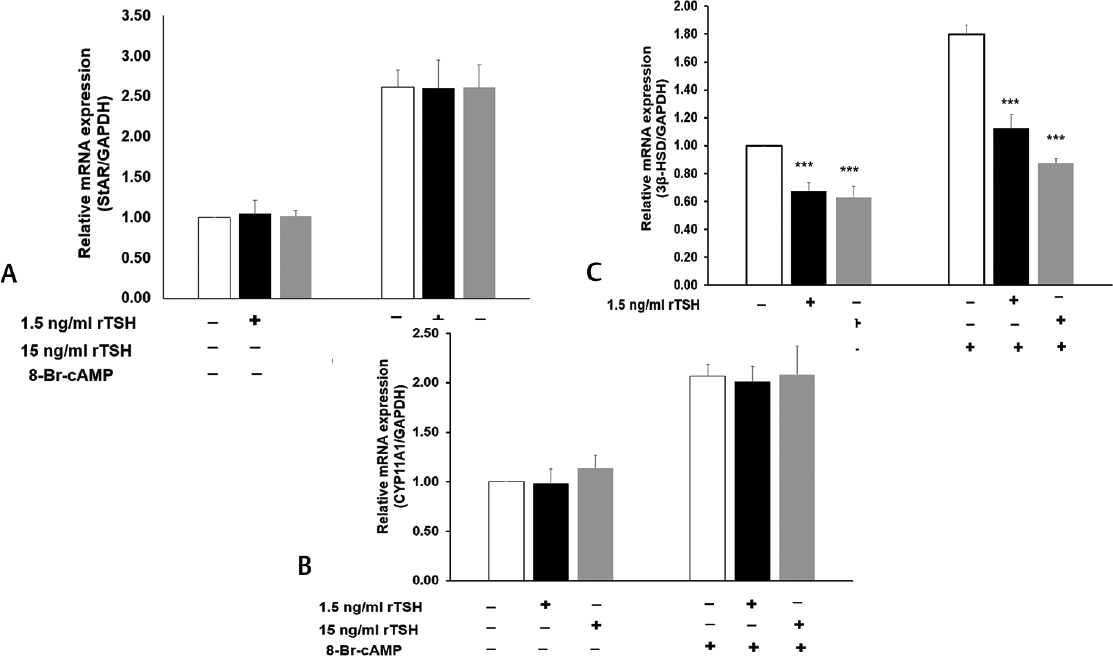
- Effect of rTSH on (A) StAR (B) CYP11A1 and (C) 3β-HSD mRNA levels by quantitative real time PCR after normalization with GAPDH. Bars represent mean ± standard deviation of three experiments; each experiment was done in triplicates (n = 9). ***indicate p < 0.001 in cells treated without and with TSH in the corresponding groups. rTSH, recombinant thyroid stimulating hormone;) StAR, steroidogenic acute regulatory protein.
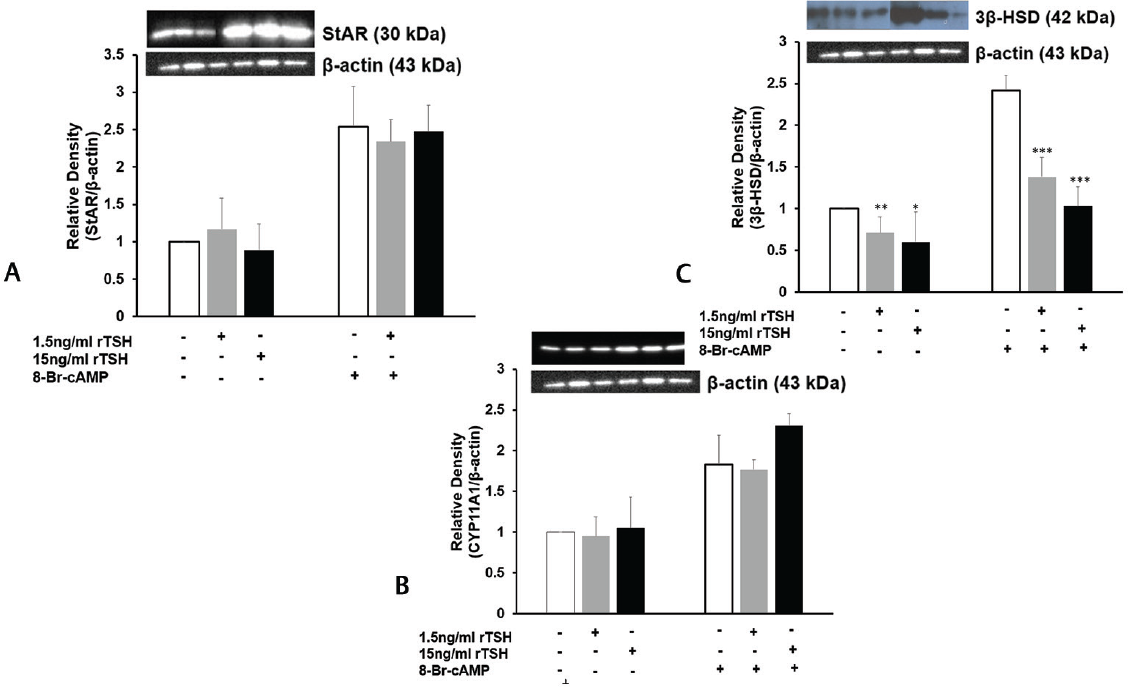
- Effect of rTSH on (A) StAR (B) CYP11A1 and (C) 3β-HSD protein levels by Western blot after normalization with β-actin. Bars represent mean ± standard deviation of densitometric measurements of Western blot bands. *indicate p < 0.05; **indicate p < 0.01; ***indicate p < 0.001 in cells treated without and with TSH in the corresponding groups. StAR, steroidogenic acute regulatory protein.
TSHR is Expressed by Mouse Leydig Tumor Cell Line-1 Cells
Next we investigated whether the action of TSH on MLTC-1 cells was mediated through its specific G-protein coupled plasma membrane receptor. We performed RT-PCR using specific primers for TSHR and western blot using anti-TSHR antibody to detect the presence of TSHR mRNA and protein in MLTC-1 cells. RNA or protein isolated from thyroid tissue was used as positive control while Vero cells, a monkey kidney derived cell line, were used as negative control. TSHR mRNA and the corresponding protein were identified in the MLTC-1 cells (►Fig. 6).
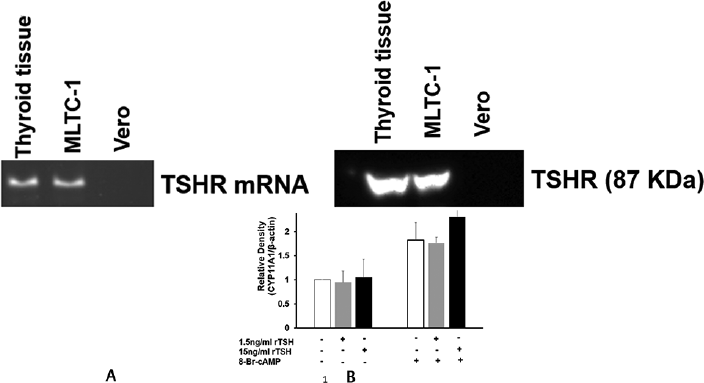
- Presence of (A) TSHR mRNA and (B) TSHR protein in thyroid tissue and mouse Leydig tumor cell line-1 cells.
Discussion
Primary Leydig cells are available only in small numbers and are often contaminated with other cell types. Therefore, for the present investigation, we have used Leydig cell-derived cell line, MLTC-1.
In Leydig cell-derived MLTC-1 cell line, TSH isolated from porcine pituitary significantly decreased basal progesterone and testosterone secretion in a dose-dependent manner (►Fig. 1A, B). TSH also inhibited LH and 8-Br-cAMP-stimulated progesterone and testosterone secretion suggesting that the inhibitory effect of TSH on steroids is distal to adenylate cyclase enzyme effect. Similarly, in primary cultures of rat granulosa cells, TSH was shown to inhibit steroid secretion.16 This finding was also confirmed in monkey and human granulosa cells.17
To rule out if the inhibitory effect of TSH on steroid secretion is nonspecific in nature, we used FSH and ACTH. FSH is structurally related to TSH; both having a common α subunit. FSH is secreted by anterior pituitary gonadotrophs. The inhibitory action of TSH on steroid secretion was not mimicked by FSH (►Fig. 2A, B). ACTH, another protein secreted by the anterior pituitary, but structurally unrelated to TSH also showed no significant effect on basal or 8-Br-cAMP-stimulated steroid secretion (►Fig. 2C, D). This suggests that the inhibitory effect of TSH on steroid secretion is unique and a specific property of the TSH hormone.
To work out the molecular mechanism of inhibition, we have used rTSH for further experiments. Pituitary preparations of TSH are mostly tainted with other contaminants. Therefore, to rule out the possibility that the inhibitory effect of TSH on steroids was not due to the contaminating molecules, we used rTSH. The rTSH, too, significantly inhibited progesterone and testosterone secretion from MLTC-1 cells confirming a direct inhibitory effect of TSH on steroid production (►Fig. 3A, B).
To investigate the inhibitory mechanism of TSH on steroid synthesis, we studied the effect of rTSH on various steroidogenic enzymes. StAR, a de novo-synthesized labile protein, catalyzes the intermitochondrial cholesterol transport.18-20 The cholesterol in the inner mitochondrial membrane is converted to pregnenolone with the help of CYP11A1 enzyme. StAR and CYP11A1 catalyze the nonsteroidogenic and steroidogenic rate-limiting steps of steroid synthesis, respectively.21,22 TSH showed no significant effect on StAR and CYP11A1 mRNA and protein levels in MLTC-1 cells (►Figs. 4 and 5). This suggests that TSH acts at a site distal to pregnenolone synthesis. However, as TSH inhibits progesterone secretion, therefore its site of action is proximal to progesterone synthesis.
In rodents, steroidogenesis is primarily through the Δ4 pathway where conversion of pregnenolone to progesterone is catalyzed by 3β-HSD enzyme.23 In mouse, the 3β-HSD1 isoform is primarily expressed in the gonads.24 Therefore, we studied the effect of rTSH on 3β-HSD1 mRNA and protein levels. In MLTC-1 cells, TSH significantly inhibited basal and 8-Br-cAMP-treated 3β-HSD mRNA and protein levels (►Figs. 4 and 5). The inhibition of 3β-HSD by TSH in this study is similar to that observed in primary cultures of rat granulosa cells.16
Next we investigated whether the direct action of TSH on MLTC-1 cells was mediated through its specific G-protein coupled plasma membrane receptor. Till date, there is only one additional report showing the presence of TSH receptors on Leydig cells that was reported after our findings were presented.25,26 However, TSH receptors have been identified on the granulosa cells of several species including rat and monkey.17,27 In MLTC-1 cells like that seen in thyroid tissue, both the TSHR and the corresponding protein were identified suggesting that the inhibitory action of TSH on steroidogenesis could be mediated through its receptor. TSHR was not identified in Vero cells, a monkey kidney derived line but has been identified in rat Leydig cells.25
However, in human thyroid tissue, when TSH binds to its receptor, it activates two G-proteins: Gs and Gq/11 28. The active Gs induce the adenylyl cyclase-cAMP pathway. The active Gq/11 activates the phospholipase C-IP3 (inositoltriphosphate)/diacyl glycerol (DAG) pathway. DAG activates the protein kinase C (PKC).28,29 In Y1 adrenocortical cells, PKC was shown to inhibit basal steroidogenesis by inhibiting the mRNA expression of two steroidogenic enzymes: P450–11 β-hydroxylase and 3β-HSD.30 In hamster cell lines, PKC has also been shown to influence the genetic expression of CYP11B2.31 The signaling mechanism by which TSH inhibits the 3β-HSD enzyme is yet to be determined in Leydig cells.
Our study is the first of its kind demonstrating a direct role of TSH on Leydig cell line and has demonstrated a molecular basis for its effects. We are also the first to demonstrate the presence of TSH receptors in Leydig cell line. There is a high probability of this physiology in human Leydig cells, but further studies are needed on primary human Leydig cell lines to redemonstrate these findings and establish the presence of this phenomenon in, as well as the TSH and these receptors in human Leydig cells.
Note
This study was previously presented as a poster presentation at the Endocrine Society meeting at Orlando, United States in April, 2017.26
Conflict of Interest
None declared.
Funding
None.
References
- Assessment of mechanisms of thyroid hormone action in mouse Leydig cells: regulation of the steroidogenic acute regulatory protein, steroidogenesis, and luteinizing hormone receptor function. Endocrinology. 2001;142(01):319-331.
- [CrossRef] [PubMed] [Google Scholar]
- Molecular mechanisms of thyroid hormone-stimulated steroidogenesis in mouse leydig tumor cells . Involvement of the steroidogenic acute regulatory (StAR) protein. J Biol Chem. 1999;274(09):5909-5918.
- [CrossRef] [PubMed] [Google Scholar]
- T3 directly stimulates basal and modulates LH induced testosterone and oestradiol production by rat Leydig cells in vitro. Endocr J. 2000;47(04):417-428.
- [CrossRef] [PubMed] [Google Scholar]
- Secretion of testicular steroids and gonadotrophins in hypothyroidism. Andrologia. 2007;39(06):253-260.
- [CrossRef] [PubMed] [Google Scholar]
- Development of the adult-type Leydig cell population in the rat is affected by neonatal thyroid hormone levels. Biol Reprod. 1998;59(02):344-350.
- [CrossRef] [PubMed] [Google Scholar]
- Increased proliferation of Leydig cells induced by neonatal hypothyroidism in the rat. J Androl. 1996;17(03):231-238.
- [CrossRef] [PubMed] [Google Scholar]
- Severe and mild neonatal hypothyroidism mediate opposite effects on Leydig cells of rats. Thyroid. 2002;12(01):13-18.
- [CrossRef] [PubMed] [Google Scholar]
- Effect of thyroid hormone on the development and gene expression of hormone receptors in rat testes in vivo. J Endocrinol Invest. 2003;26(05):435-443.
- [CrossRef] [PubMed] [Google Scholar]
- Hypoandrogenaemia is associated with subclinical hypothyroidism in men. Int J Androl. 2007;30(01):14-20.
- [CrossRef] [PubMed] [Google Scholar]
- Leydig cells, thyroid hormones and steroidogenesis. Indian J Exp Biol. 2005;43(11):939-962.
- [Google Scholar]
- Establishment of gonadotropin-responsive murine leydig tumor cell line. J Cell Biol. 1982;94(01):70-76.
- [CrossRef] [PubMed] [Google Scholar]
- Detection of P450c17-independent pathways for dehydroepiandrosterone (DHEA) biosynthesis in brain glial tumor cells. Proc Natl Acad Sci U S A. 1998;95(06):2862-2867.
- [CrossRef] [PubMed] [Google Scholar]
- Leydig cell re-generation and expression of cell signaling molecules in the germ cell-free testis. Reproduction. 2008;135(06):851-858.
- [CrossRef] [PubMed] [Google Scholar]
- Defining thyrotropin-dependent and -independent steps of thyroid hormone synthesis by using thyrotropin receptor-null mice. Proc Natl Acad Sci U S A. 2002;99(24):15776-15781.
- [CrossRef] [PubMed] [Google Scholar]
- Effects of partial silencing of genes coding for enzymes involved in glycolysis and tricarboxylic acid cycle on the enterance of human fibroblasts to the S phase. BMC Cell Biol. 2015;16:16.
- [CrossRef] [PubMed] [Google Scholar]
- Thyroid stimulating hormone inhibits rat granulosa cell steroidogenesis in primary culture. Endocrine. 1995;3(03):215-220.
- [CrossRef] [PubMed] [Google Scholar]
- Thyroid stimulating hormone (TSH) receptor on granulosa cells. Fertil Steril. 2011;96(03):S118.
- [CrossRef] [Google Scholar]
- The purification, cloning, and expression of a novel luteinizing hormone-induced mitochondrial protein in MA-10 mouse Leydig tumor cells . Characterization of the steroidogenic acute regulatory protein (StAR) J Biol Chem. 1994;269(45):28314-28322.
- [CrossRef] [PubMed] [Google Scholar]
- The orphan nuclear receptors steroidogenic factor-1 and liver receptor homolog-1: structure, regulation, and essential roles in mammalian reproduction. Physiol Rev. 2019;99(02):1249-1279.
- [CrossRef] [PubMed] [Google Scholar]
- Leydig cells: formation, function, and regulation. Biol Reprod. 2018;99(01):101-111.
- [CrossRef] [PubMed] [Google Scholar]
- Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr Metab (Lond). 2010;7:47.
- [CrossRef] [PubMed] [Google Scholar]
- Steroid hormones: synthesis, secretion, and transport. In: Belfiore A, LeRoith D, eds. Principles of Endocrinology and Hormone Action. Cham: Springer International Publishing; 2016. p. :1-31.
- [CrossRef] [Google Scholar]
- Testicular steroid synthesis: organization and regulation. In: Knobil E, Neill J, eds. The Physiology of Reproduction (2nd). New York, NY: Raven Press; 1994. p. :1335-1362.
- [Google Scholar]
- The multiple murine 3 beta-hydroxysteroid dehydrogenase isoforms: structure, function, and tissue- and developmentally specific expression. Steroids. 1997;62(01):169-175.
- [CrossRef] [PubMed] [Google Scholar]
- Effects of hyper- and hypothyroidism on the development and proliferation of testicular cells in prepubertal rats. Anim Sci J. 2017;88(12):1943-1954.
- [CrossRef] [PubMed] [Google Scholar]
- A unique anti-gonadotropic effect of TSH on Leydig cell derived Mltc-1 Line. The Endocrine Society Meeting. Available at: https://endo.confex.com/endo/2017endo/meetingapp.cgi/Paper/31402. (accessed )
- [Google Scholar]
- Thyrostimulin, but not thyroid-stimulating hormone (TSH), acts as a paracrine regulator to activate the TSH receptor in mammalian ovary. J Biol Chem. 2010;285(06):3758-3765.
- [CrossRef] [PubMed] [Google Scholar]
- The human thyrotropin receptor activates G-proteins Gs and Gq/11. J Biol Chem. 1994;269(19):13733-13735.
- [CrossRef] [PubMed] [Google Scholar]
- The thyroid and its regulation by the TSHR: evolution, development, and congenital defects. In: Luster M, Duntas LH, Wartofsky L, eds. The Thyroid and Its Diseases: A Comprehensive Guide for the Clinician. Cham: Springer International Publishing; 2019. p. :219-233.
- [CrossRef] [Google Scholar]
- Protein kinase C is a tonic negative regulator of steroidogenesis and steroid hydroxylase gene expression in Y1 adrenal cells and functions independently of protein kinase A. Mol Endocrinol. 1993;7(08):1021-1030.
- [CrossRef] [PubMed] [Google Scholar]
- Control of CYP11B2 gene expression through differential regulation of its promoter by atypical and conventional protein kinase C isoforms. J Biol Chem. 2001;276(11):8021-8028.
- [CrossRef] [PubMed] [Google Scholar]



